Home | Archive | AJA @ Nature | Online Submission | News & Events | Subscribe | APFA | Society | Links | Contact Us | 中文版 |
|
Epidermal growth factor receptor pathway substrate 8 (Eps8) expression in maturing testis
Alain Wunsch, Kai Strothmann, Manuela Simoni, Jörg
Gromoll, Eberhard Nieschlag, C. Marc Luetjens
Institute of Reproductive Medicine of the University, D-48129 Mnster,
Germany
Asian J Androl 2004 Sep; 6: 195-203
Keywords: gene expression; isoform; Sertoli cell; gonocyte
Abstract
Aim: Although epidermal growth factor receptors are expressed in the testes, whether they signal through epidermal growth factor receptor pathway substrate 8 (Eps8) is unknown. Here we evaluated the expression pattern of Eps8 in the maturing testis. Methods: The expression of Eps8 was analysed by Northern blotting, immunocytochemistry and Western blotting in primary Sertoli cell cultures and in testicular tissue of rodents. Results: Eps8 is specifically expressed in gonocytes, Leydig and Sertoli cells of the neonatal rats and in Leydig and Sertoli cells of the adult rats and mice. Although gonocytes express Eps8, no signal was found in prepubertal or mature spermatogonia and the expression level of Eps8 in Sertoli cells increases with age. No regulation of Eps8 expression in primary immature rat Sertoli cells by Follicle stimulating hormone (FSH) was detected by Western blotting. Conclusion: Eps8 seems to be involved in the growth factor-controlled regulation of cell proliferation and differentiation in the seminiferous epithelium. Eps8 is a possible marker for gonocytes and in Sertoli cells it could be involved in crosstalk with other growth factor pathways.
1 Introduction
Sertoli cells play an important role in mammalian spermatogenesis by creating a unique microenvironment for the developing germ cells. The epidermal growth factor receptor (EGFR) is mainly localized in the immature and adult Sertoli and Leydig cells of rodent and primate testis [1-3]. The distinct function of the EGFR pathway in Sertoli cells is still under debate due to the fact that the EGFR "knockout" mouse shows no defect on testis growth, indicating that the pathway must be compensated [4]. Conversely, inhibition of the EGFR tyrosine kinase does have a growth inhibition effect on the testis [4], demonstrating that this pathway is of importance for the mitogenic activity of Sertoli cells. Moreover, over expression of epidermal growth factor leads to hypospermatogenesis in mice [5].
A recent investigation in immature rat Sertoli cells using cDNA arrays led to the identification of factors induced or repressed by the Follicle stimulating hormone (FSH) applied to the cell culture for up to 24 hours [6].
Using a murine SK-11 cell line originally described by Walther et al [7], we previously established two new Sertoli cell lines stably expressing the wild type FSH receptors (FSHR) or the constitutively active Asp567Gly FSHR. Gene expression profiling of these cell lines was performed using commercially available mouse cDNA arrays and led to the identification of Eps8 as a factor expressed in these Sertoli cells [8]. The constitutively active FSHR cells showed a down-regulation of Eps8 mRNA transcripts. However, no data on the protein expression level is known so far.
Eps8 exists in two protein isoforms, p97eps8 and p68eps8, and is involved in the growth and differentiation regulatory pathways of different cell types [9]. Due to a number of binding domains, Eps8 is seen as a multivalent molecular link assembling intracellular transducers with receptor tyrosine kinases (RTKs) and possibly also establishing connections with pathways activated by G protein-coupled receptors (GPCRs) [10]. In particular, Eps8 participates in actin remodelling through Rac and in inhibiting RTK internalisation through Rab5, thereby prolonging receptor signaling [11]. Since Eps8 is believed to play an important role in the control of proliferation-differentiation processes [12, 13], we further analysed the expression pattern of Eps8 in maturing rodent testis and primary Sertoli cells.
2 Materials and methods
2.1 Sertoli cell line
The SK-11 cells [7] were grown at 33 (5 % CO2) in Dulbecco's modified Eagle medium (Gibco) containing 10 % FCS (PAA Laboratories, Cölbe, Germany). The total RNA was isolated from these cells as described below.
2.2 Primary Sertoli cell culture
Animal experiments were performed according to the German Law on Animal Care, Protection and Experimentation. Male Sprague-Dawley rats (Charles River, Sulzfeld, Germany), 10 days old, were sacrificed by decapitation after CO2 anaesthesia. The primary Sertoli cells were isolated by sequential enzymatic digestion as described previously [14, 15]. They were then cultured in serum-free medium (DMEM containing 1000 mg/mL glucose from Gibco) containing 2 % penicillin-streptomycin and 1 % non-essential amino acids (Gibco) for 4 days at 34 (5 % CO2) on plastic Petri dishes (for Western blot analysis) or on glass coverslips (for immunocytochemistry). Stimulation with FSH for different time periods was performed in serum-free culture medium by the addition of purified urinary FSH (1 IU/mL) (Fertinorm HP, Serono, Unterschleißheim, Germany). At the end of the incubation, cells were harvested with RIPA buffer (TRIS 25 mmol/L pH7.5, sodium orthovana-date 2 mmol/L, EGTA 0.5 mmol/L, NaF 10 mmol/L, sodium pyrophosphate 10 mmol/L, -glycerophosphate 80 mmol/L, NaCl 25 mmol/L, Triton 100 1 %, sodium deoxycholate, SDS 0.05 %, protease inhibitor (Sigma P8340 0.1 %) for subsequent Western analysis or fixed in 4 % paraformaldehyde for immunocytochemistry.
2.3 Testis tissue
Testis tissue was obtained from 3 different litters of mice (CD1) and Sprague-Dawley rats on day 5, 10, 20 and 60, respectively. For the immunohistochemistry, rat testis of day 5, 10, 20 and 60 as well as from mice on day 60 was fixed in Bouin's solution and sectioned. For Northern blotting, murine testis of postnatal day 5, 10 and 60 were snap frozen.
2.4 Northern blotting
Total RNA was isolated from SK-11 cells and postnatal mouse testes (day 10) by a modified Chomczynski and Sacchi phenol-chloroform extraction method using UltraspecTM (Biotecx Laboratories, Inc., Houston, USA). Total RNA 20 g was separated on 1 % agarose-formaldehyde gel, checked for integrity under UV-light and transferred to NybondTM nylon membranes (Amersham Pharmacia, Freiburg, Germany). Blots were probed with a gel-purified [-32P]-dATP-labelled Eps8 cDNA fragment (primers: forward-primer: 5'-AAC TGT GAC CGC AAC GGC AAC TTC C-3' reverse-primer: 5'-ATC CTG TAG GGC ACA TCC TAT AAA CA-3' amplifying a 311bp cDNA) and exposed to Phosphorimager screens (Molecular Dynamics, Freiburg, Germany). Eps8 mRNA signals were quantified using ImageQuant software (Molecular Dynamics) and normalised against -actin. All Northern blots were repeated twice with different samples.
2.5 Western blotting
Protein concentrations in cell homogenates of murine SK-11 cells and rat primary Sertoli cells (postnatal day 10) in RIPA buffer were determined by the BCA-protein assay (reagents from Pierce, Rockford, IL, USA). After loading the 10 % polyacrylamide gels with 10 g protein per pocket, electrophoresis was run at 200 V and transferred at 100 mA for 1h to the Trans-Blot® nitrocellulose membranes (Bio-Rad, Mnchen, Germany). After overnight blocking with 5 % milk powder in 1PBS/0.01 %, Tween 20 blots were incubated with a monoclonal anti-Eps8 primary Ab (1:5000; Transduction Laboratories, Heidelberg, Germany). Enhanced chemiluminescent detection was performed using the ECL plus kit by Amersham and signals were quantified using the Molecular Dynamics ImageQuant software. All Western blottings were performed 3 times.
2.6 Immunocytochemistry
Cells were washed (1PBS),
fixed (paraformaldehyde 4 % v/v) and permeabilised with ice-cold 0.1 %
Triton x-100 in PBS. After washing and blocking with
5 % horse serum in 0.3 % Triton x-100/PBS, cells were incubated for 2
h with the monoclonal anti-Eps8 primary Ab (1:500; Transduction Laboratories)
and a rabbit polyclonal antibody specific for
a-smooth muscle actin diluted 1:50 (Sigma A-2547, USA) followed by washing.
Detection was done with an anti-mouse biotin conjugated Ab (Sigma B1404/1:1000)
in 1 % horse serum in 0.3 % Triton x-100/PBS and Streptavidin Texas red
(1:1000) and an anti-rabbit conjugated with FITC 1:100 (Sigma F-0382)
in PBS. Nuclear staining was performed with Hoechst 33258 (1 mg/mL in
PBS). Fluorescence was observed using an upright Axioskop microscope (Zeiss,
Oberkochen, Germany). Digital images of equal exposure were acquired using
an AxioCam camera and AxioVision software (Zeiss). The detection of Eps8
and a-smooth muscle actin was performed in duplicates.
2.7 Immunohistochemistry
Bouin-fixed and paraffin-embedded testis sections of adult mice were stained with a polyclonal rabbit antibody against Eps8 (generous gift of Dr. P. P. di Fiore, Milano, Italy). Due to the fact that the monoclonal Eps8 antibody did not work on Bouin-fixed tissue and the poly-clonal Eps8 antibody showed a strong background in the Western blotting, we had to use two different antibodies to detect Eps8 at the protein level. After deparaffination, blocking of endogenous peroxidases with 0.3 % H2O2 and 1 h incubation with 1 % BSA and 5 % horse serum, antigen retrieval was performed in a microwave oven at 80 for 10 min. Sections were incubated with the Eps8 antibody (1:200) for 1 h followed by incubation with a peroxidase-conjugated anti-rabbit antibody (Sigma 0-9169, 1:400) for 1 h. After washing, DAKO-EnVision (DAKO Diagnostika, Germany) was added for 30 min, followed by washing and incubation with Newfuchsin (DAKO Diagnostika) for 30 min. In a final step, sections were counterstained with hematoxylin for 10 sec and mounted under cover slips with Dako Faramount (DAKO Diagnostika) prior to observation using an upright microscope (Axioskop, Zeiss, Oberkochen, Germany) at different magnifications (objectives 10, 25, 40 and 100. Digital images of equal exposure were acquired with a CCD camera (AxioCam) controlled by the image software (AxioVision).
3 Results
Two different Eps8 transcripts were demonstrated by Northern blotting in the Sertoli cell line SK-11 (Figure 1A), where the larger transcript was more abundant. In the immature mouse testicular RNA preparations, only the longer Eps8 transcript was found. By Western blotting, two Eps8 protein isoforms could be detected in SK-11 cells: a larger isoform of ~ 97 kDa (p97Eps8) and a more abundant, smaller isoform of ~ 68 kDa (p68Eps8) (Figure 1B).
Figure 1. Expression of mRNA Eps8 expression in Sertoli cells. A: Eps8 mRNA expression in SK-11 cell lines and immature mouse testis. Eps8 mRNA expression is apparently expressed stronger in SK-11 compared to whole testis (~2.5-fold). B: Western Blot analysis of Eps8 expression in SK-11 Sertoli cell line. Both isoforms (p68Eps8 and p97Eps8 expression) could be found in SK-11 cell line and p68Eps8 expression was relatively higher as suggested by the Northern blot data.
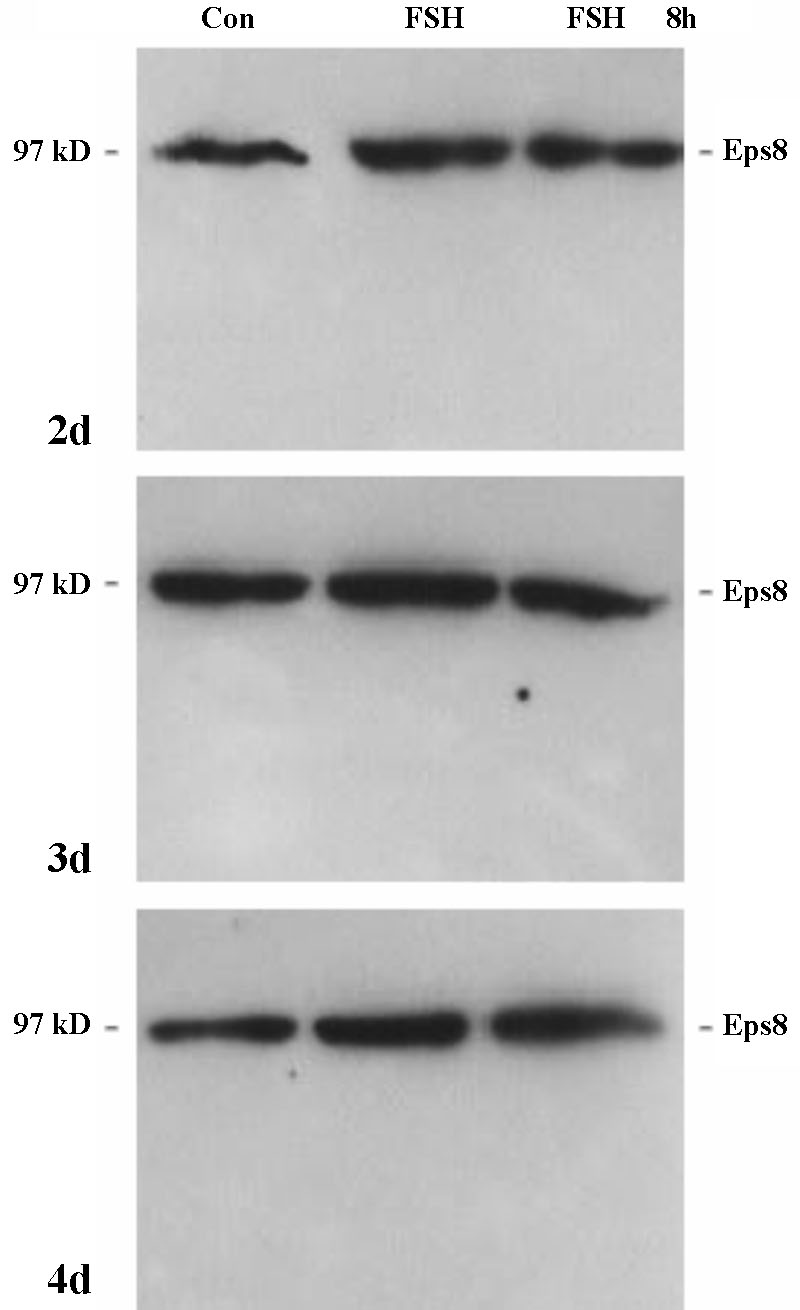
In order to analyse whether Eps8 expression in Sertoli cells is modulated by FSH, we isolated and cultured Sertoli cells from 10-day-old rats. As shown in Figure 2, cultured Sertoli cells stained weakly positive for Eps8 and grew in clusters surrounded by a few peritubular cells as described earlier [14]. We estimated that our cultures consisted of about 90 % Sertoli cells and that peritubular cells did not overgrow up to at least 5 days in culture in the presence of FSH, as shown by specific peritubular cell staining with an anti smooth muscle actin antibody [14] (Figure 2, lower panel). The expression of Eps8 and its putative regulation by FSH were further analysed by Western blotting (Figure 3). Sertoli cells were cultured for different time periods in the presence and absence of FSH. Under basal conditions, i.e. in the absence of FSH stimulation, Eps8 expression was constant over time for up to 8 days in culture (not shown). Stimulation with FSH (1 IU/mL) for 2, 4, 24 and 48 hours did not result in significant changes in Eps8 expression (48 hours shown in Figure 3). In all repeated experiments (n=3) when the Sertoli cells were cultured in the presence of FSH (8 h/day) for up to 4 days the normalised expression of p97Eps8 was constant. In the primary Sertoli cells culture, only the p97Eps8 expression was found (Figure 3) with no effect by FSH treatment.
{kind=link}
Figure 2. Primary culture of rat Sertoli cells with some peritubular cells. Sertoli cells are stained for Eps8 and peritubular cells for their specific cell marker -smooth muscle actin. The nuclei are stained with Hoechst. Scale bar =100 m. The histogram is a quantification of Sertoli cells compared with the total amount of cells in cell culture. Data are representative from n = 4 cell cultures in two separate experiments.
Figure 3. Western Blot analysis of Eps8 expression in primary rat Sertoli cells after 2, 3 and 4 days of culture in the absence (con) or presence of FSH (1 IU/mL). The FSH stimulation was constant (FSH) or restricted to 8 hours/day (FSH 8 h). p97eps8 expression amount remained unchanged during culture in the presence of FSH given for 8 h/day for 2, 3 and 4 days.
To further analyse the in vivo expression of Eps8, immunohistochemical analyses were performed. Testicular tissue from rats and mice of different ages (postnatal, prepubertal and adult) showed specific staining for Eps8 protein in the seminiferous tubule and the interstitium, according to a pattern which changes with the age of the animals (Figure 4 and Figure 5). In the seminiferous tubule Eps8 staining was particularly intense in the gonocytes and spermatogonia in neonatal rats (5 to 10 days of age) and was otherwise absent from other germ cell types at all ages investigated (Figure 4). The gonocytes of postnatal day 5 express Eps8, but not the Sertoli cells already organising the seminiferous epithelia. The Sertoli cells of postnatal day 20 rats were also not stained, but Eps8 expression in mature Sertoli cells was evident in adult animals (60 days) both in rats and mice (Figure 5). Of the interstitial cells only the fetal Leydig cells showed Eps8 staining (postnatal day 10 and 20). On postnatal day 60, expression of Eps8 was always apparent in the interstitial cells of rodent testes and the Leydig cells showed especially intense staining. Neither the endothelial nor the peritubular cells were positive for the Eps8 signal at all time points investigated.
Figure 4. Immunohistochemical localisation of Eps8 expression in prepubertal rat testis. Left panels: with first antibody against Eps8; right panels: without first antibody. In testis of 5-day-old rats several gonocytes (*) and spermatogonia (o) express Eps8. After 10 days expression level strongly decreased in these two cell types, whereas fetal Leydig cells (arrows) start expressing the protein. On postnatal day 20 only maturing to adult type Leydig cells express Eps8. Scale bar =100 m.
Figure 5. Immunohistochemical detection of Eps8 in adult rat and mouse testis. In adult testis Sertoli cells express specifically Eps8 (black arrows). The long cell extensions of the Sertoli cells (black arrow) reaching into the centre of tubuli are clearly visible, demonstrating the localisation of Eps8 in close proximity with spermatids (bottom right image). Leydig cells are also positive for Eps8 (green arrows). Bottom left demonstrates a control section leaving out the primary antibody with no staining. Upper panels scale bar =100 m. Middle and bottom left scale bar =25 m. Bottom right scale bar=50 m.
As an additional confirmation, Eps8 mRNA was shown to be highly expressed in the prepubertal testis and the expression level decreased progressively with age (Figure 6). These results, together with those of immunohistochemistry, suggest that the decline in the Eps8 signal from prepubertal to adult testis observed by Northern blotting (Figure 6) is due to the increasing numbers of maturing germ cells within the seminiferous epithelium that do not express Eps8.
Figure 6. Eps8 mRNA expression in testis of 5, 10 and 60 day-old mice (normalised against -actin) and densitometric quantification (AU: arbitrary densitometric units). Testicular Eps8 mRNA content decreases from the prepubertal to the adult animal.
4 Discussion
In this paper we describe the expression of Eps8 in the testis and, in particular, the expression pattern decreasing and disappearing in the germ cells and appearing in the Sertoli cells of the matured tissue.
Eps8 is a substrate for the EGFR and several other RTKs [7], which was originally implicated in the routing of EGF-dependent mitogenic signals. In their original study, Fazioli et al [9] showed that Eps8 is functionally and physically coupled to the EGFR and that the overexpression of Eps8 in different target cells increases their mitogenic responsiveness to EGF. While Eps8 acts as a positive regulator of mitogenesis and is induced in response to growth-promoting stimuli (e.g. EGF, serum or phorbol esters), its expression (but not that of other EGFR substrates) is strongly down-regulated during terminal differentiation of non-proliferating murine myogenic cells [12]. The localisation of Eps8 in the Leydig and Sertoli cells of the testis is not surprising since Eps8 expression is ubiquitous [10] and it is well known that EGF and other RTKs are expressed and affect the function of testicular somatic cells [2, 16]. However, unexpectedly we found that the expression pattern of Eps8 changes from mitogenic cells, the germ cells, which continuously divide prior to differentiation, to cells which are known not to proliferate after puberty. Eps8 might be one of the very few markers for gonocytes and very early spermatogonia and might be slowly down-regulated after a transition time point, which has been described for mice on day 3 and for rats on day 5 [17, 18]. Only very few spermatogonia were still positive for Eps8 on day 10 and no germ cell staining was detected in older testis. In the prespermatogenesis period, male germ cells (gonocytes) begin to reproliferate and move to the basement membrane of the seminiferous tubule. Although these two events - reproliferation and relocation - are important for the establishment of spermatogenesis, they have not been analysed in great detail. However, these events fit very well with the time point of Eps8 down-regulation in gonocytes, suggesting a connection of the biochemical changes and the morphological changes of the germ cell line. For an improvement of stem cell culture and to follow their differentiation pattern to meiotic competent germline cells, such a marker for gonocytes will have tremendous importance [19].
According to prior descriptions of EGFR expression [20], we found that the interstitial cells and gonocytes were positively stained for Eps8 on postnatal day 5. Interestingly, Eps8 disappears progressively from germ cells after the early neonatal period and appears to be solely localised in the Leydig and Sertoli cells in the adult testis. It is possible that the Eps8 expression in Sertoli cells described here is part of the developmental switch in the expression of the TGFa/EGFR/Eps8 signalling system occurring in the postnatal phase and at puberty.
Eps8 has been shown to exist in two isoforms, p97Eps8 and p68Eps8 [9]. The antibodies employed in this study are designed to recognise both isoforms and we detected both isoform bands of 97 and 68 kDa. In the mouse Sertoli cell line SK-11, the p68Eps8 isoform is expressed at higher levels than the p97Eps8 isoform. The p68Eps8 isoform is probably synthesised from an alternatively spliced mRNA [9]. A variable distribution and expression in response to stimuli of these two isoforms depending on the cell line and organism investigated has already been observed by several authors [10, 12]. Whether the two isoforms exert different functions is presently not known.
In primary rat Sertoli cells, the expression of Eps8 was detected by immunocytochemistry and Western blot analyses and remained constant after a prolonged treatment with FSH. In our experimental setting only the p97Eps8 isoform was expressed in the primary Sertoli cells. To avoid excessive desensitisation and down-regulation of the FSHR induced by prolonged FSH treatment, cells were cultured with FSH 8 h/day [21]. In this way we attempted to mimic the in vivo situation, where Sertoli cells are exposed over a long term to changing concentrations of FSH. No effect of FSH on Eps8 expression was found, suggesting that Eps8 does not require FSH to transduce a differentiating signal in Sertoli cells. In a previous study, results of Northern blotting in the SK-11/MUT cell line suggested that Eps8 is regulated by the constitutively active FSHR [8], however, our protein expression results show that no down-regulation of the Eps8 protein takes place and the quantity of Eps8 mRNA is not correlated with the protein quantity. This might be due to the fact that only a limited amount of mRNA might be enough to maintain the protein expression level as shown for other mRNAs. However, the in vitro situation with immortalised cells with a chronic stimulus can not be directly compared to the primary Sertoli cell situation. Transgenic mice overexpressing a constitutively active FSHR in their Sertoli cells might be useful to analyse further this aspect [22]. In the primary culture cells the proliferative Sertoli cell index did not change significantly over seven days of culture (Figure 5). The 97 kDa isoform has been shown to be expressed in growing cells [12], which could explain the discrepancy between the Eps8 expression in differentiated Sertoli cells in vivo and proliferating cells in vitro.
In conclusion, our data identify Eps8 as a novel marker of gonocytes. The transition of gonocytes to spermatogonia is accompanied by a down-regulation of Eps8 in gonocytes and an up-regulation in Sertoli cells. This germ cell transition is correlated with the mitotic active phase to populate the testis tubules with spermatogonia. Adult Sertoli cells are mitotically arrested and express Eps8 compared to mitotically active immature Sertoli cells. It will be interesting to learn more details about the functional role of Eps8 in Sertoli cells and the signal transduction network regulating its expression.
Acknowledgements
This work was supported by a grant from the Deutsche Forschungsgemeinschaft (Confocal Research Group "The Male Gamete: Production, Maturation, Function" grant FOR 197-3). We are grateful to Dr. P.P. di Fiore, European Institute of Oncology, Milano, Italy, for the Eps8 antiserum. The excellent technical assistance of R. Sandhowe-Klaverkamp and J. Salzig and the language editing by S. Nieschlag, M.A., are gratefully acknowledged.
References
[1] Suarez-Quian
CA, Dai MZ, Onoda M, Kriss RM, Dym M. Epidermal growth factor localization
in the rat and monkey testis. Biol Reprod 1989; 41: 921-32.
[2] Suarez-Quian CA, Niklinski W. Immunocytochemical localization of the
epidermal growth factor receptor in mouse testis. Biol Reprod 1990; 43:
1087-97.
[3] Radhakrishnan B, Suarez-Quian CA. Characterization of epidermal growth
factor receptor in testis, epididymis and vas deferens of non-human primates.
J Reprod Fertil 1992; 96: 13-23.
[4] Levine E, Cupp AS, Miyashiro L, Skinner MK. Role of the transforming
growth factor-alpha and the epidermal growth factor receptor in embryonic
rat testis development. Biol Reprod 2000; 62: 477-90.
[5] Wong RW, Kwan RW, Mak PH, Mak KK, Sham MH, Chan SY. Overexpression
of epidermal growth factor induced hypospermatogenesis in transgenic mice.
J Biol Chem 2000; 275: 18297-301.
[6] McLean DJ, Friel PJ, Pouchnik D, Griswold MD. Oligonucleotide microarray
analysis of gene expression in follicle-stimulating hormone-treated rat
sertoli cells. Mol Endocrinol 2002; 16: 2780-92.
[7] Walther N, Jansen M, Ergn S, Kascheike B,
Ivell R. Sertoli cell lines established from H-2Kb-tsA58 transgenic mice
differentially regulate the expression of cell-specific genes. Exp Cell
Res 1996; 225: 411-21.
[8] Strothmann K, Simoni M, Mathur P, Siakhamary S, Nieschlag E, Gromoll
J. Gene expression profiling of mouse Sertoli cell lines. Cell Tissue
Res 2004; 315: 249-57.
[9] Fazioli F, Minichiello L, Matoska V, Castagnino P, Miki T, Wong WT,
et al. Eps8, a substrate for the epidermal growth factor receptor
kinase, enhances EGF-dependent mitogenic signals. EMBO J 1993; 12: 3799-808.
[10] Wong WT, Carlomagno F, Druck T, Barletta C, Croce CM, Huebner K,
et al. Evolutionary conservation of the EPS8 gene and its mapping
to human chromosome 12q23-q24. Oncogene 1994; 9: 3057-61.
[11] Di Fiore PP, Scita G. Eps8 in the midst of GTPases. Int J Biochem
Cell Biol 2002; 34: 1178-83.
[12] Gallo R, Provenzano C, Carbone R, Di Fiore PP, Castellani L, Falcone
G, et al. Regulation of the tyrosine kinase substrate Eps8 expression
by growth factors, v-Src and terminal differentiation. Oncogene 1997;
15: 1929-36.
[13] Maa MC, Hsieh CY, Leu TH. Overexpression of p97Eps8 leads to cellular
transformation: implication of pleckstrin homology domain in p97Eps8-mediated
ERK activation. Oncogene 2001; 20: 106-12.
[14] Simoni M, Schuhmann B, Weinbauer GF. In vitro Sertoli cell
bioassay of follicle-stimulating hormone (FSH)-serum from different animal
species alters the morphology of rat Sertoli cells without affecting their
response to FSH. Gen Comp Endocrinol 1994; 95: 99-108.
[15] Schlatt S, de Kretser DM, Loveland KL. Discriminative analysis of
rat Sertoli and peritubular cells and their proliferation in vitro:
evidence for follicle-stimulating hormone-mediated contact inhibition
of Sertoli cell mitosis. Biol Reprod 1996; 55: 227-35.
[16] Spiteri-Grech J, Nieschlag E. Paracrine factors relevant to the regulation
of spermatogenesis - a review. J Reprod Fertil 1993; 98: 1-14.
[17] de Rooij DG, Russell LD. All you wanted to know about spermatogonia
but were afraid to ask. J Androl 2000; 21: 776-98.
[18] Nagano R, Tabata S, Nakanishi Y, Ohsako S, Kurohmaru M, Hayashi Y.
Reproliferation and relocation of mouse male germ cells (gonocytes) during
prespermatogenesis. Anat Rec 2000; 258: 210-20.
[19] Ohbo K, Yoshida S, Ohmura M, Ohneda O, Ogawa T, Tsuchiya H, et
al. Identification and characterization of stem cells in prepubertal
spermatogenesis in mice. Dev Biol 2003; 258: 209-25.
[20] Cupp AS, Skinner MK. Expression, action and regulation of transforming
growth factor alpha and epidermal growth factor receptor during embryonic
and perinatal rat testis development. J Androl 2001; 22: 1019-29.
[21] Themmen AP, Blok LJ, Post M, Baarends WM, Hoogerbrugge JW, Parmentier
M, et al. Follitropin receptor down-regulation involves a cAMP-dependent
post-transcriptional decrease of receptor mRNA expression. Mol Cell Endocrinol
1991; 78: R7-13.
[22] Nordhoff V, Gromoll J, Foppiani L, Luetjens CM, Schlatt S, Kostova
E, et al. Targeted expression of human FSH receptor Asp567Gly mutant
mRNA in testis of transgenic mice: role of human FSH receptor promoter.
Asian J Androl 2003; 5: 267-75.
Correspondence to:
Prof. E. Nieschlag, FRCP, Institute
of Reproductive Medicine of the University, Domagkstr. 11, D - 48129 Mnster,
Germany.
Tel: +49-251-835 6097, Fax: +49-251-835 6093
Email: nieschl@uni-muenster.de
Present address of Kai Strothmann: Schering AG, 13342 Berlin, Germany.
A. Wunsch and K. Strothmann contributed equally to the study
Received 2003-12-02 Accepted 2004-04-28
